Samuel P. Reed, Alejandro A. Royo, Walter P. Carson, Castilleja F. Olmsted, Lee E. Frelich, Peter B. Reich
{"title":"Multiple disturbances, multiple legacies: Fire, canopy gaps and deer jointly change the forest seed bank","authors":"Samuel P. Reed, Alejandro A. Royo, Walter P. Carson, Castilleja F. Olmsted, Lee E. Frelich, Peter B. Reich","doi":"10.1111/1365-2745.14459","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 2","pages":"353-370"},"PeriodicalIF":5.6000,"publicationDate":"2024-12-11","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14459","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14459","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
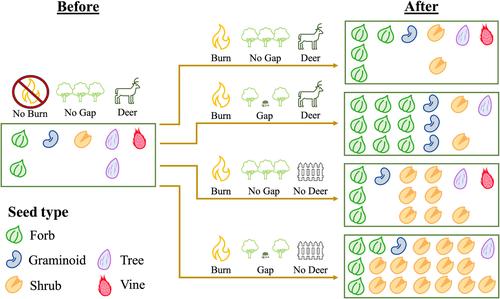
随着土地利用的改变、气候变化或土著居民的被迫迁移,世界各地的森林经历了历史性干扰制度的重大变化(Bowman et al., 2011;Gilliam, 2016;Gotmark, 2013;Kelly et al., 2023)。北美、欧洲和亚洲的许多中温带森林已经变得更加均匀,经历了严重的灭火,有蹄动物的浏览也增加了(Carpio et al., 2021;Frelich, 2002;Hai et al., 2023;McDowell et al., 2020;Pascual-Rico等人,2021)。这些变化的条件为共同进化并依赖于历史干扰模式的植物物种创造了不利的环境,例如全球分散的橡树(Quercus)物种(Carrero等人,2020;Tinner et al., 2005)。随着干扰制度的改变,植物组成的变化导致管理者恢复或操纵干扰,以支持生物多样性和生态系统功能(Long, 2009;Stanturf et al., 2014)。然而,我们对多种历史干扰的重新引入如何影响生物多样性的理解是新生的,这代表了我们在温带森林系统的长期管理和恢复方面的关键知识差距。北美森林是一个广阔的生态系统,在过去的一个世纪里,它们的干扰机制经历了巨大的变化(Abrams, 2005;Hanberry,Nowacki, 2016;Vander Yacht et al., 2020;韦伯斯特等人,2018)。这种情况在阿巴拉契亚阔叶林中尤为严重,这些阔叶林已经失去了橡树(栎属)树木的再生,并正在过渡到更湿润的、以枫为主导的(宏属)系统。艾布拉姆斯,2008;Pile Knapp et al., 2024)。这种从橡树林到枫树林的过渡是由土著居民被迫搬迁和他们使用文化焚烧作为管理工具而开始的(Abrams等人,2021;Pile Knapp等,2024;波勒斯,2015)。随后是19世纪末和20世纪初的大规模森林砍伐和大面积野火(Lafon et al., 2017)。对这些野火的负面看法导致了一个世纪以来国家批准的火灾排除和抑制,这有利于枫树的生长和更湿润的林下植被(Alexander等人,2021;Arthur et al., 2021)。结果,阿巴拉契亚森林以年龄均匀的林分为主,很少有大中型林分(直径15米;175平方米)树冠间隙和罕见的低强度火灾(Clebsch &;用校车接送学生,1989;Nowacki,艾布拉姆斯,2008;Raymond et al., 2009)。在阿巴拉契亚地区,火灾的复发间隔现在超过了1万年,而不是土著管理下的历史上1到20年的火灾复发间隔和闪电点燃的火灾(Lafon et al., 2017)。与此同时,北美东部大部分地区的白尾鹿(Odocoileus virginianus)种群数量大幅增加,高于历史基线(高于4至8只/平方公里),这取决于它们的种群密度,推动了生态变化,类似于欧洲和亚洲许多其他地区鹿群过多的影响(Côté等人,2004;Iijima等,2023;Reed et al., 2022;Valente et al., 2020)。为了扭转历史管理的长尾效应并维持以橡树为主的植物群落,森林管理者正在重新引入干扰,如规定的焚烧,通过采伐树木创造树冠间隙,以及通过狩猎或将脆弱地区围起来降低鹿的密度(Nuttle等人,2013;Raymond et al., 2009)。重新引入多重干扰可以成为恢复和指导生态群落内部变化的有力工具(Abrams等人,1985;Batllori等人,2019;Reed et al., 2023;Sasaki et al., 2015;Yantes et al., 2023)。例如,在北美和欧洲,低强度的火灾和树冠间隙的形成相结合可以促进橡树的生长,而这些干扰本身的效果较差(Brose et al., 2013;Hutchinson et al., 2024;Izbicki et al., 2020;peterson et al., 2020)。在这个例子中,幸存的橡树代表了干扰后的遗产,其大致特征是在干扰后保留在景观上的适应性、个体和生物量(Cuddington, 2011;富兰克林等人,2000)。干扰遗产可以是物质的(例如木材和营养池)和信息的(例如物种的适应反应和遗传物质),尽管这两类并不是相互排斥的(Johnstone等人,2016)。在给定区域发生的每次干扰都会改变先前干扰的遗留群落,在某些情况下,干扰组合和时间可能会导致独特的群落,这取决于所讨论的干扰如何相互作用(Anoszko等人,2022)。 因此,在美国东部森林和世界各地的温带森林中,与这些干扰单独的遗产相比,低强度火灾、冠层间隙创造和有蹄类动物的觅食组合的干扰遗产可能在决定森林如何重组和发展到未来方面具有特别重要的作用(Cuddington, 2011;Seidl et al., 2014;特纳和Seidl, 2023)。为此目的,土壤种子库是一个重要但研究不足的实体,它可能受到重新引入的干扰的强烈影响,并可能影响未来的干扰制度(Archibold, 1979;Ferrandis et al., 1996;摩根,Neuenschwander, 1988;Pakeman,小,2005;苏萨,1984)。种子库是一种生殖适应,允许植物以休眠种子的形式在地下持续存在,其中土壤可以作为地上干扰的缓冲(Baskin &;巴斯金,2022;汤普森,1987)。森林种子库已被证明是世界各地温带森林生物多样性的储存库,拥有许多草本和木本早期演替物种(Grubb et al., 2013;Plue et al., 2010;Yang等人,2021)。种子库也是遗传多样性的潜在来源(Levin, 1990;McCauley, 2014),使种子库成为物质和信息遗产。在扰动中幸存下来的发芽植物最终成熟并释放种子,重新建立种子库过程,使植物群落能够在未来的扰动中重组,从而根据返回土壤的种子建立另一种遗产(Baltzer等人,2021;Faliń平方公里列阵,1999;Grubb, 1988;凯悦酒店,卡斯珀,2000;Seidl,特纳,2022)。假设,更多的干扰将导致种子库更类似于地上植被,因为草本层是均匀的,少数野生物种存活和繁殖(Ma et al., 2021)。种子库的这些变化和干扰会产生长期的生态后果。例如,在19世纪末和20世纪初,美国的木材采伐和森林大火可能使灌木红莓传播并使其长寿的种子饱和森林种子库,在整个美国东部受到植被干扰后,形成了长达一个世纪的红莓重再生遗产(Dunn et al., 1982;彼得森,卡森,1996)。然后,红莓可以作为一种顽强的林下植物存活数十年(Donoso &;Nyland, 2006;Kern等人,2012)。规定的燃烧、树冠间隙的创造和鹿的浏览都为种子库中的新植被生长和种子库的变化提供了独特而重要的机会(Gioria等人,2022;Muscolo et al., 2014;舒勒,2010)。规定的火清除植物材料,通过增加光、热、烟和营养来催化种子发芽(Keeley &;Fotheringham, 2000;Ooi 2012;Pausas et al., 2022)。Pausas和Lamont(2022)发现,在全世界容易发生火灾的生态系统中,约42%的种子库物种适应在高温或烟雾中发芽。林冠间隙增加了林下资源,如光照、土壤温度和土壤湿度,这些对种子发芽至关重要(Dalling &;布朗,2009;Pakeman,小,2005)。当植物发芽时,火隙和冠层间隙都会导致种子库中的种子暂时枯竭,但随着时间的推移,新建立的植被会生长、繁殖并补充种子库(Auld &;德纳姆,2006;信田,赤坂,2020)。这种补充过程可能会被有蹄类食草动物破坏,因为长期的过度浏览会限制种子的结子,减少植物的丰度,并通过将组成转移到浏览耐受物种而降低长期的林下植物多样性(Brody &;欧文,2012;Pendergast et al., 2016)。这些直接消耗效应可能间接降低种子库物种的丰度和多样性(Beauchamp et al., 2013;(2019)。然而,在鹿群数量较少且与历史估计相似的地区,鹿的啃食已被证明通过减少原本具有竞争力的野生物种的丰度来增加林下多样性,这可能导致更多样化的种子库(Royo等人,2010;Yacucci等人,2024)。尽管在美国东部和更广泛的温带系统中,对重新引入的干扰的实验测试越来越普遍和同时发生(Kleinman等人,2019;托姆和Seidl, 2016),我们对个体和组合低强度火灾,树冠间隙和有蹄类食草动物如何改变长期森林种子库的理解是最小的。这凸显了我们对干扰后遗产的理解存在重大差距,因为种子库对于在干扰的情况下维持森林生物多样性至关重要。 因此,指导我们研究的主要问题是:与每个单独的干扰相比,多个重新引入的干扰是否会对种子库造成更大的长期变化?为了验证这个问题,我们使用了一个独特的多干扰森林实验,通过控制燃烧来控制低强度火灾,通过围篱和除草剂注射来制造树冠间隙,通过围栏封闭来控制鹿的密度。实验开始十三年后,我们对每种干扰组合处理下的种子库进行采样,并在多个时间点与现有植被进行比较,测试了不同干扰处理下种子组成的变化情况。我们预计低强度火灾将是种子密度和多样性增加的主要驱动因素,因为阿巴拉契亚生态系统历史上经历了频繁的低强度火灾,许多植物物种可能受到火灾的青睐(H1)。同样,我们预计冠层间隙会导致种子库密度和多样性的适度增加,这与Royo等人(2010)(H2)指出的冠层间隙增加的地上植物多样性相一致。我们假设林火与林冠间隙结合会导致种子库密度和多样性的最大增加,从而导致种子群落组成的变化(H3)。基于鹿的食草性对地上植物生长和繁殖的负面影响的研究,我们预计鹿对种子库密度和多样性有负面影响,特别是在与火结合的情况下(H4)。最后,当将种子库与现存植被进行比较时,我们预计种子库群落与高度受干扰地块的现存植被最相似,因为许多种子库物种受到干扰,可能已经饱和了种子库(H5)。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
