A pan-European citizen science study shows population size, climate and land use are related to biased morph ratios in the heterostylous plant Primula veris
Tsipe Aavik, Triin Reitalu, Marianne Kivastik, Iris Reinula, Sabrina Träger, Evelyn Uuemaa, Marta Barberis, Arjen Biere, Sílvia Castro, Sara A. O. Cousins, Anikó Csecserits, Eleftherios Dariotis, Živa Fišer, Grzegorz Grzejszczak, Cuong Nguyen Huu, Kertu Hool, Hans Jacquemyn, Margaux Julien, Marcin Klisz, Alexander Kmoch, Nikos Krigas, Attila Lengyel, Michael Lenhard, Desalew M. Moges, Zuzana Münzbergová, Ülo Niinemets, Baudewijn Odé, Hana Pánková, Meelis Pärtel, Ricarda Pätsch, Theodora Petanidou, Jan Plue, Radosław Puchałka, Froukje Rienks, Ioulietta Samartza, Julie K. Sheard, Bojana Stojanova, Joachim P. Töpper, Georgios Tsoktouridis, Spas Uzunov, Martin Zobel
{"title":"A pan-European citizen science study shows population size, climate and land use are related to biased morph ratios in the heterostylous plant Primula veris","authors":"Tsipe Aavik, Triin Reitalu, Marianne Kivastik, Iris Reinula, Sabrina Träger, Evelyn Uuemaa, Marta Barberis, Arjen Biere, Sílvia Castro, Sara A. O. Cousins, Anikó Csecserits, Eleftherios Dariotis, Živa Fišer, Grzegorz Grzejszczak, Cuong Nguyen Huu, Kertu Hool, Hans Jacquemyn, Margaux Julien, Marcin Klisz, Alexander Kmoch, Nikos Krigas, Attila Lengyel, Michael Lenhard, Desalew M. Moges, Zuzana Münzbergová, Ülo Niinemets, Baudewijn Odé, Hana Pánková, Meelis Pärtel, Ricarda Pätsch, Theodora Petanidou, Jan Plue, Radosław Puchałka, Froukje Rienks, Ioulietta Samartza, Julie K. Sheard, Bojana Stojanova, Joachim P. Töpper, Georgios Tsoktouridis, Spas Uzunov, Martin Zobel","doi":"10.1111/1365-2745.14477","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"114 1","pages":""},"PeriodicalIF":5.6000,"publicationDate":"2025-01-22","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://besjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14477","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14477","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
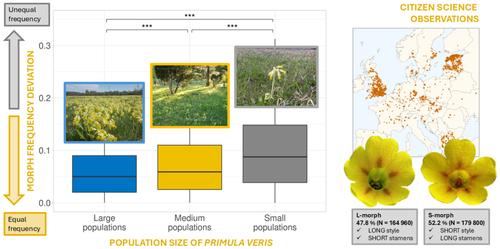
由于全球土地利用变化,上个世纪出现了前所未有的栖息地丧失和破碎化(Haddad et al., 2015)。随着气候变化,这一趋势对生物多样性的几个方面产生不利影响,造成遗传多样性的丧失(Des Roches et al., 2021;Laikre et al., 2020;Schlaepfer等人,2018)、物种丰富度(Tilman等人,2017)和相关生态系统服务(Cardinale等人,2012)。然而,并非所有物种对这些因素的反应都相同,有些物种比其他物种更容易受到上述威胁。在植物中,某些物种的反应可能取决于它们的生活史、功能性状、表型可塑性和生物地理起源(De Kort et al., 2021;Hamrick,Godt, 1996)。此外,破碎化和气候变化对植物共生伙伴的影响,如传粉者(Bennett et al., 2020;Rodger等人,2021)、种子传播者(Donoso等人,2022)或菌根真菌(Kiesewetter等人,2023;Outhwaite et al., 2022;Senapathi et al., 2017),可能会影响植物物种对全球变化因素的相对脆弱性。由于生境丧失和气候变化对传粉者的潜在负面影响,异交、动物传粉植物可能比无性繁殖、自交或风媒植物更容易受到气候变化和栖息地破碎化的影响(Aguilar et al., 2008;Bennett et al., 2020;Rodger et al., 2021)。此外,传粉媒介丰度和多样性的减少可能最终导致植物-传粉媒介网络的变化(Zoller等人,2023),可能引发异交或自交不亲和减少的表型选择(Bodbyl Roels &;凯利,2011;Cheptou, 2021;Jacquemyn et al., 2012;Opedal, 2019)。然而,我们对当代景观中生殖植物性状如何响应气候变化和土地利用变化的理解仍然有限(Pontarp et al., 2023)。授粉不足对那些具有阻碍自花授粉特性的植物物种造成了特别的威胁,例如异种花柱。异质花柱是一种由遗传决定的花的多态性,表达在雌性和雄性生殖器官的相互定位中(Barrett, 2019)。它已经在至少28个植物科中独立进化(Barrett, 2019)。异花柱植物的种群包括两种(二花柱种)或三种(三花柱种)的花柱和花药长度互反的变体。形态之间的差异也可能表现在柱头乳突和花粉粒的大小和形态上(Costa, Castro, et al., 2017)。花多态性的研究历史悠久,查尔斯·达尔文是第一个提出这种互惠的花多态性有助于确保植物个体之间的异交(Barrett &;海岸,2008;达尔文,1862;Simón-Porcar et al., 2022)。最近的研究表明,异花柱植物物种通常以遗传决定的不相容系统为特征,促进了非分类交配(Costa, Ferrero等,2017;Huu et al., 2016, 2022;Keller et al., 2014)。异花柱植物的自交不亲和性和相关性状,包括花药位置、花柱高度、花粉形态之间的大小差异以及生化自交不亲和性由位于S座的一组紧密相连的基因控制,在二花柱植物物种中,S座在一种形态(S-morph)中是半合子的,而在另一种形态(L-morph)中则不存在(Huu et al., 2016)。不同的形态在种群中通常具有平衡的形态频率(等密度)。然而,剧烈的景观变化和植物种群大小的下降可能导致等多态的随机偏差(Endels et al., 2002;ksamry et al., 2003),甚至导致一种(二花科植物)或两种变种(三花科植物;费列罗等人,2020;Heuch, 1980)。不平衡的形态比会减少异种植物种群中合适交配伴侣的数量、生殖产量和遗传多样性(Kaldra et al., 2023;ksamry等人,2003;Meeus et al., 2012)。由于非同种交配和对传粉者的强烈依赖,异花柱植物因此特别容易受到栖息地丧失的负面影响(Brys等,2004),这可能最终导致自亲和性增加,并在花粉严重稀缺的条件下导致异花柱的破裂(Wang等,2020)。此外,气候条件的变化可能对植物与传粉者的相互作用产生重大影响。温度和降水的变化,以及极端天气事件频率的增加,都可能导致植物-传粉媒介网络的物候解耦,改变传粉媒介的觅食模式,增加花粉降解和更强的花蜜稀释(Lawson &;兰德,2019;Settele et al., 2016)。 在异花柱物种中,降水模式变化引起的较高湿度可导致不同形态花粉的不同生存力(Aronne et al., 2020)。因此,土地利用和气候变化的综合影响可能会对具有复杂交配系统的植物造成更大的威胁,例如异花柱(Aronne et al., 2020;Stefanaki et al., 2015;Thomann et al., 2013)。报春花(Primula veris L.)是花柱异质性研究中常用的二花丛模式种(如ksamry et al., 2003;Nowak et al., 2015;Potente et al., 2022)。这种草本植物生长在欧亚大陆的农村和山区(特别是南欧),它们的首选栖息地是半自然草原和半开放森林。然而,欧洲90%的半天然草地面积已经消失(Dengler et al., 2020),对许多草地物种的发生和遗传多样性产生不利影响(Kiviniemi, 2008;Lienert, 2004;Lindborg et al., 2005),包括P. veris (Brys &;Jacquemyn, 2009;Kery et al., 2000;Van Rossum et al., 2004)。在平衡状态下,由于负频率依赖的平衡选择,两种形态的比例相等(Heuch, 1979)。与等密度的偏差与狐尾假蝇种群规模的下降有关(Aavik等人,2020;Kaldra et al., 2023;ksamry et al., 2003)。由于一种或另一种变体的相对损失是完全随机的,因此,这种频率相等的变体的随机偏差预计不会影响种群中变体的一般比例。最近的一项研究在其北部分布极限(爱沙尼亚)的一千多个种群中探索了狐尾草的形态比例,结果表明,短型s型总体上超过长型l型,s型占主导地位的种群也超过了s型(Aavik等人,2020)。这一发现表明,其他确定性过程塑造了变异的偏差,导致一种特定的变异类型比另一种非随机流行。先前的研究发现,其他报春花物种在其分布范围边缘附近的种群中,形态间和形态内相容性和自交性存在相当大的差异(Shao et al., 2019;Van Daele等人,2024;Zhang et al., 2021)或沿海拔梯度(Yuan et al., 2017)。当殖民化事件导致小而孤立的种群时,这种交配系统的破坏就会发生,在这种情况下,自相容性是有利的。传粉媒介的稀缺可能进一步促进了这种转变,有利于采用自交和同质作为确保成功繁殖的策略(Yuan等人,2017)。最近的证据表明,这些变化可能是S位点相关基因突变或重排的结果(Mora-Carrera et al., 2023)。除了传粉媒介短缺的自然原因外,人类引起的土地利用变化减少了传粉媒介的丰度,并可能导致对传粉媒介依赖性降低的转变(Brys &;Jacquemyn, 2012;Cheptou, 2021)。尽管已经证明花粉限制对植物繁殖策略的影响及其不利的适应性后果(Rodger et al., 2021),但在人类活动强烈改变的景观中,植物交配系统的变化很少被研究(Cheptou, 2021)。为了填补关于异花柱植物形态比例大尺度模式的知识空白,我们在其大部分欧洲分布范围内探索了羊尾草的种群内形态频率。我们在28个欧洲国家发起了一项公民科学运动,使欧洲假单胞菌种群形态比率的大规模数据收集成为可能。特别是,我们的目标是检验以下假设:较小的种群规模可能导致变异与平衡频率的显著偏差(ksamry et al., 2003)。我们假设这些偏差将是随机的,即不支持一种或另一种形态。在草地和林地生境有限的景观中,即耕地和建成区比例较高的同质景观中,由于生境破碎化,预计其偏离等积性的程度会更大。这些影响可能是随机的(独立于一种特定的形态)或确定性的,即有利于一种特定的形态(Aavik等人,2020),通过景观组成对传粉者的影响来调节,正如对其他二花物种的研究所表明的那样(Shao等人,2019;Yuan等,2017;Zhang等人,2021)。或者,如有限的证据所显示的,P. veris中L-morphs的部分形态内相容性(Brys &;Jacquemyn, 2015;Wedderburn,Richards, 1990),可能导致L-morphs在低生境可利用性的景观中优于S-morphs。 在这种条件下,相对于具有几乎完全的形态内兼容性的s型,l型具有相对较多的合适交配伴侣,这将使这种形态类型具有优势。然而,只有在P. elation (Van Daele et al., 2024)中才证明了另一种可能性,即s型异型在碎片化种群中可能会被过度代表,这是由于进化出的较低的两性杂交、减少的生理自交不亲和和花粉从高花药落到低柱头上的更自主的自花授粉的综合影响。相对形态频率,无论是随机的还是确定的,都可能受到气候因素的影响,如降水和温度,这些因素可能影响植物与传粉者的相互作用,因为传粉者在高温和多雨条件下不那么活跃(Ganuza et al., 2022;Jiao等,2023;Kammerer et al., 2021;劳森,兰德,2019;Teixido et al., 2022)。此外,由于风速增加对传粉者活动的负面影响,植物与传粉者的相互作用可能被破坏(Hennessy et al., 2021)。我们假设,在风速较高的地区,种群将表现出更大的偏离相同形态频率(等plethy),因为风媒介的花粉传播或传粉者有限导致的传粉服务中断可能会不同地影响两种形态的繁殖成功率。最后,我们预计,由于人类对欧洲农村景观的压力增加,以及由此导致的传粉媒介丰富度和丰度的下降,我们将发现异花柱的同质个体,这表明异花柱的破坏或破坏。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.






 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
