Clément Vallé, Isabelle Le Viol, Jean Nabias, Karine Princé, Frédéric Gosselin
{"title":"Tree species identity shapes the relationship between canopy cover and herb-layer species in temperate forests","authors":"Clément Vallé, Isabelle Le Viol, Jean Nabias, Karine Princé, Frédéric Gosselin","doi":"10.1111/1365-2745.14479","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 3","pages":"582-597"},"PeriodicalIF":5.6000,"publicationDate":"2025-02-04","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14479","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14479","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
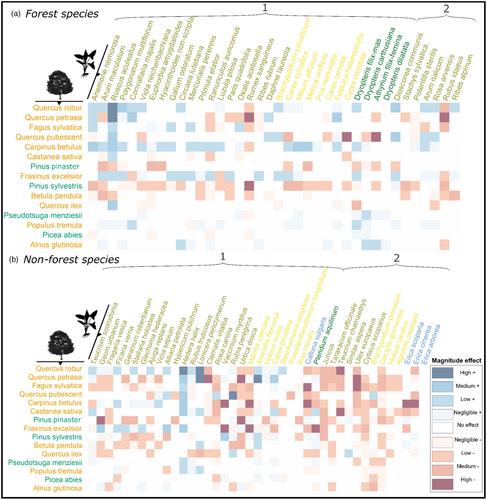
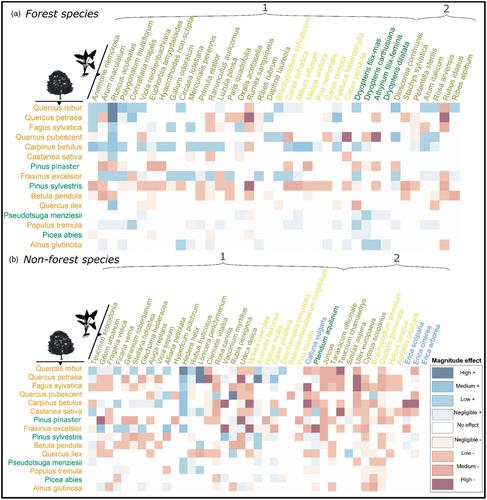
在森林生态系统中,林下植被代表了植物生物多样性的很大一部分,高达90% (Gilliam &;罗伯茨,2014;Thrippleton等人,2016),并提供多种生态系统服务,如养分循环、碳固存和能量流(Landuyt等人,2019;Mölder等人,2008;穆勒,2014;Wang等人,2021)。它不仅为哺乳动物和昆虫等物种群体提供栖息地,还塑造了其他物种的分布(Fayt et al., 2006;马歇尔,Hawthorne, 2012),而且通过与发芽幼苗的资源竞争来调节树木的再生(Coomes &;Grubb, 2000;Gilliam, 2007)或促进过程(例如,通过菌根网络的小气候缓冲或交换;卡拉威,沃克,1997;Zilliox,Gosselin, 2014)。因此,掌握草本层分布的驱动因素对于指导可持续森林管理和生物多样性保护计划至关重要(Köhl et al., 2020)。草本层群落对不同尺度的土地利用历史、非生物和生物因子等多种因素做出响应(Barbier et al., 2008;Ellenberg和Leuschner, 2010;Gilliam, 2007;Hermy,Verheyen, 2007)。气候影响其广泛分布和时间变化。在局部尺度上,森林地面适宜微环境的有效性是一个强大的驱动因素(Kelemen et al., 2014),包括土壤成分和条件、水和光的有效性或小气候温度(Depauw, 2020;魏等人,2020)。然而,这些因素部分取决于森林结构和树木组成(Augusto et al., 2003)。例如,树冠可以拦截99%的入射光(Coomes &;Grubb, 2000),激烈竞争地下资源(Coomes &;Grubb, 2000;德国等人,2017)和调节小气候(Gottschall等人,2019;Lembrechts et al., 2019)。由于营养循环、降雨分配、遮荫能力和树叶透射光质量的差异,树木也可以与特定的草本物种联系在一起(Barbier, Balandier, &;Gosselin, 2009;Rawlik et al., 2018)。树种特性也会导致凋落物组成(如化感化合物)和积累,影响草本层物种的萌发和表现(Bertin et al., 2003;Rodríguez-Calcerrada et al., 2011)。因此,众所周知,各种上层组成特征会影响下层草本群落,但它们对单个物种分布的影响程度仍然存在问题(Ampoorter et al., 2015;Barbier et al., 2008;德国等人,2021;Gilliam,罗伯茨,2014)。森林树种群落的分布、多样性和结构在很大程度上受到管理实践的影响(Tinya et al., 2021),管理实践也会影响草本层,例如边缘管理(Govaert et al., 2020)。树种多样性不仅可以增加环境异质性(资源异质性假说,Ricklefs, 1977),还可以通过非加性树种丰富度效应创造新的环境条件(Ball et al., 2008),从而对草本层的对应物产生积极影响。然而,最近的研究发现物种丰富度效应的结果并不一致(Ampoorter et al., 2015;Both et al., 2011;passion et al., 2018)和树种识别(Coppi et al., 2019;Corcket et al., 2020;german et al., 2021)或树种丰度(Barbier, Chevalier, et al., 2009)可能是草本层组成、生物量和系统发育多样性的更重要驱动因素。这可以解释为,考虑树种身份比分类物种丰富度更能直接整合它们的功能差异,使其更能预测影响草本层物种分布的潜在理论生态机制。树木身份可以更好地预测每个草本物种生长的潜在非生物限制因子(即李比希限制因子定律,Austin, 2007),并通过更准确地描述冠层树木的功能互补性(即资源异质性假说)更好地捕获潜在的生态位异质性(因为不同树种捕获的资源份额差异很大)。此外,如果某些树种对草本层施加更大的竞争压力(如高营养或光吸收树种),这可能导致草本层物种的建立更适应抗逆性,而不是适应竞争或干扰(Grime的CSR策略;污垢,2006)。因此,树木的某些功能分组也可能与预测冠层对草本层物种的影响更相关。 例如,许多针叶树种倾向于积累凋落物,比大多数阔叶树更能酸化土壤(Augusto et al., 2015;Díaz-Calafat等人,2023),有利于耐酸草本物种,限制核心林草本物种的发芽和存活(thomas等人,2013)。然而,Barbier等人(2008)警告说,不要简化针叶树和阔叶树对草本物种的影响,特别是因为一些物种不具有在其类群中主要存在的相同特征(例如,山毛榉阔叶树的厚凋落物会使土壤酸化)。此外,基于树木身份的替代功能分组可能与捕获该地层对草本物种的影响更相关(例如,Raunkiaer生命形式类型,演替阶段或通过竞争获取基质对土壤酸度或湿度的Ellenberg亲和力)。树木的Raunkiaer生命形式通过调节光照、水分和营养物质来决定草本层的物种组成(Gilliam &;罗伯茨,2014;马奎尔和Forman, 1983)和创建特定的微型网站(Glick &;Matlack, 2021),例如较高树木的倾斜丘(导致特定的草本层物种组成;Spicer et al., 2018)。此外,树木演代(或动态)阶段可以直接影响草本物种的组成和数量(Rybar等,2023)(例如,通过透射光的质量和数量的变化),也可以间接影响影响该草本层的生物的丰度,例如食草昆虫(Hilmers等,2018)。尽管如此,Barbier等人(2008)建议将树-林下相互作用作为物种特异性关系进行探索,因为树种的身份可能会通过多维功能特征和机制的组合影响草本层物种,而这些特征并未被所考虑的特征完全捕获。由于数据集的要求,例如足够的分辨率和高样本量,物种关联的检索具有挑战性(vall<e:1>等人,2023)。专注于树种身份的研究往往受到样本量小和空间范围有限的限制,导致只鉴定了少数物种特异性关系(berg<s:1> et al., 2017;Dölle等人,2017;Nagel et al., 2019)。探索层与层之间关系的潜在机制的实验研究可能非常耗时,并且需要充分的设计(Both et al., 2011)。因此,确定大量树木和草本物种之间的潜在联系至关重要。几位作者(Gilliam et al., 1995;Peet et al., 2014)还假设,上层-下层联动可能只发生在一定时期(约20年的林分发育)之后,因为这两层主要(且主要)在早期发育过程中对不同的非生物因素做出反应(即树木的光有效性和草本层物种的水和养分有效性;Gilliam et al., 1995)。此外,随着时间的推移,由于树木倒下等事件,新的环境条件被创造出来的可能性会增加(巴特尔斯&;陈,2010)。此外,树冠对叶片生长的封闭导致树冠空间中物种之间的互补树冠结构(Jucker等人,2018;Williams et al., 2017)。这些发现可以解释通过实验研究难以检测树木身份对林下物种的影响(Germany et al., 2017,2021)。此外,几位作者强调缺乏积极联系的经验证据(卡尔顿和;Maycock, 1981;Ellenberg, 1988)。这强调了大型经验数据集的宝贵贡献,包括很大比例的长期建立的林分。发现树种与草本层物种之间的潜在关联,评估树种身份效应的大小,可以帮助我们了解和预测草本层的空间分布。在这项研究中,我们依赖于全国范围内的数据集耦合上层-下层采样协议,以精细分辨率描述法国大陆的树冠组成和草本层覆盖(Vidal et al., 2005)。该议定书得益于熟练的森林观察员对采样地点的详细经验环境描述(非生物和管理参数)。这增加了检测到的关联不仅仅是由于物种对分析中未考虑的因素做出类似反应的可能性(Poggiato et al., 2021)。因此,我们假设在控制树木分类物种多样性效应和已知影响草本层分布的主要可测量环境因子的情况下,可以检测草本层物种与树种覆盖身份之间的关联(H1.a)。我们同意Barbier et al.(2008)和Barbier, Chevalier等人的观点。 (2009)认为,通过“阔叶”与“针叶树”的简单二元分类,无法充分表达树木身份的影响。考虑到这两层相互连接的复杂机制,我们提出替代的树木功能分组可能提供树木对草本植物影响的更全面的表示,尽管没有分类可以完全涵盖树木身份效应的全部(H1.b)。我们还假设草本物种也可以根据其栖息地偏好对树种身份覆盖有共同的反应(H1.c)。例如,与森林草本层物种相比,非森林草本层物种对优势树种的身份或其丰度的负响应程度更高,在基于受控样地类型采样(Barbier, Chevalier, et al., 2009)或在统计模型中考虑样地类型变量(Zilliox &;Gosselin, 2014)。虽然遥感技术的进步促进了对冠层树木的监测,可以识别许多物种和许多个体,但草本层并没有直接受益于这一工具。我们的第二个假设提出,使用冠层树种身份覆盖可以利用这些物种关联改善对许多草本层物种个体分布的预测(vall<e:1>等人,2023;Wilkinson et al., 2021),在新的站点(H2)。这可以间接地利用遥感技术的进步,并代表了对这一层的理解和空间预测的重大进步,这一层的调查既困难又耗时。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.






 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
