Nathéo Beauchamp, Georges Kunstler, Laura Touzot, Paloma Ruiz-Benito, Emil Cienciala, Jonas Dahlgren, Paweł Hawryło, Matija Klopčič, Aleksi Lehtonen, Vladimír Šebeň, Jarosław Socha, Miguel A. Zavala, Benoit Courbaud
{"title":"Light competition affects how tree growth and survival respond to climate","authors":"Nathéo Beauchamp, Georges Kunstler, Laura Touzot, Paloma Ruiz-Benito, Emil Cienciala, Jonas Dahlgren, Paweł Hawryło, Matija Klopčič, Aleksi Lehtonen, Vladimír Šebeň, Jarosław Socha, Miguel A. Zavala, Benoit Courbaud","doi":"10.1111/1365-2745.14489","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 3","pages":"672-688"},"PeriodicalIF":5.6000,"publicationDate":"2025-02-06","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14489","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14489","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
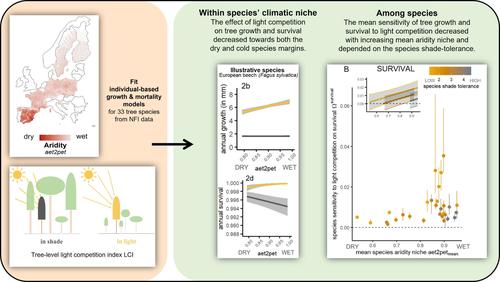
众所周知,光竞争是森林中的一个关键过程,是森林结构和树木动态的重要决定因素(Pacala et al., 1996),树种的遮荫耐受性水平差异很大(Valladares &;Niinemets, 2008)及其对光竞争的敏感性(Kunstler et al., 2011)。此外,估算树木光竞争的模型比表示其他资源(如水或养分)竞争的模型更先进,这使得在更大尺度上研究光竞争效应的变化成为可能(Craine &;Dybzinski, 2013)。这将我们带入了第二条探究线:使用涵盖广泛环境梯度的大规模研究来进一步了解森林对气候的响应(Ruiz-Benito et al., 2020)。如果我们希望包括物种的气候边缘,大规模的研究是至关重要的,在气候边缘,特定物种的人口统计表现可能会有所不同(Kunstler et al., 2021)。大规模研究的另一个主要优势是,它们使比较众多物种成为可能,这有助于回答物种之间对竞争的敏感性是否因其气候生态位或生态策略而异的问题。气候竞争相互作用的方向和强度可能会随着物种对资源限制的容忍程度而变化(Maestre等,2009)。例如,Kunstler等人(2011)发现,竞争对树木生长的重要性随着生产力的提高而降低,沿着温度和干旱的生物气候梯度,竞争对不耐阴物种的平均重要性高于耐阴物种。这些结果强调了不仅要在大地理范围内研究多物种的竞争-气候相互作用,而且要根据不同物种的生态策略比较不同物种对光竞争的敏感性的重要性。在此,我们对光竞争对整个欧洲单株树木生长和生存的影响进行了大规模研究,这得益于一个超过100万棵树木的数据库的可用性,其中包括从西班牙到斯堪的纳维亚半岛的九个欧洲国家。首先,为了分析光竞争的影响,我们从SamsaraLight光线追踪模型(Courbaud et al., 2003, 2015)中推导出树级光竞争指数,这是一个空间明确的基于树木的模型,根据林分中每棵树的3D树冠对光束的拦截和衰减来估计给定树木的光截获量。我们用树种特有的树冠异速生长来表示空间上的树冠结构。然后,我们考虑了两个气候梯度:温度和干旱。我们使用基于土壤结构、月水通量和融雪量的水平衡模型来推导样地水平的干旱指数。最后,我们拟合了基于特定树种的生长和死亡模型,作为气候和光照竞争的函数。然后,我们利用这些模型预测了不同气候和光竞争水平下的树木年生长和存活,以检验光竞争的影响是否在两种气候梯度上有所不同,在干燥气候下的高干旱和寒冷气候下的低温限制了树木的动态。主要假设是,净光竞争效应将取决于遮阳的负面影响(减少碳同化)和积极影响(减少蒸发需求、减少霜冻胁迫等)之间的平衡。根据这些气候压力的相对重要性,这种平衡可能会沿着气候梯度发生变化。此外,由于其他气候条件的限制,处于完全光照下的好处也不尽相同。当供水量较低或低温应力较高时,性能的改善应较小。我们在两个不同的尺度上解决了这个问题:(i)物种内部——在给定物种的气候边缘之间;(ii)物种之间——通过比较不同物种对不同荫凉耐受性水平和不同平均气候生态位的反应。我们假设(i)在给定物种的气候生态位中,光竞争对生长和生存的影响在寒冷或干燥的物种压力边缘处较弱;(ii)在物种之间和整个欧洲,对生长和生存的光竞争的平均敏感性对于耐荫物种来说较弱,对于平均气候生态位位于炎热干燥的地中海地区或寒冷的北方地区的物种来说。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
