Xiao-Long Li, 李孝龙, Jun Zhou, 周俊, Hong-Qiu Du, Fei Peng, Hongtao Zhong, Yanhong Wu, Ji Luo, Shouqin Sun, Yue-Xin Ming, Hongyang Sun, Yang Chen, Jun Wasaki, Hans Lambers
{"title":"Plant nutrient-acquisition strategies contribute to species replacement during primary succession","authors":"Xiao-Long Li, 李孝龙, Jun Zhou, 周俊, Hong-Qiu Du, Fei Peng, Hongtao Zhong, Yanhong Wu, Ji Luo, Shouqin Sun, Yue-Xin Ming, Hongyang Sun, Yang Chen, Jun Wasaki, Hans Lambers","doi":"10.1111/1365-2745.70017","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 4","pages":"988-1003"},"PeriodicalIF":5.6000,"publicationDate":"2025-03-03","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.70017","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.70017","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
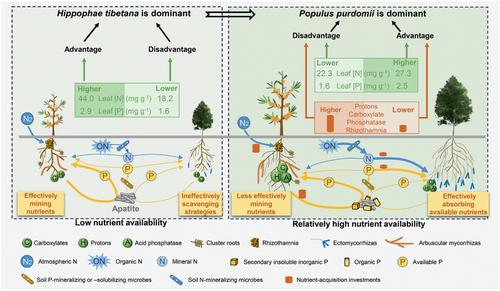
了解物种替代对揭示植物群落演替机制至关重要(Buma et al., 2017;Cantera et al., 2024),这几十年来一直是生态学的挑战。初级演替过程中的植物物种更替可能与生态过程有关,例如光竞争(Buma等人,2019)、植物特异性土壤病原体的积累(Van der Putten等人,1993)、化感作用(Chapin等人,1994)以及对不同养分有效性的响应差异(Tilman, 1985)。氮(N)和磷(P)限制通常发生在陆地生态系统中(Oldroyd &;草地,2020)。植物已经进化出一系列养分获取策略(NAS),以响应土壤养分的不同可用性和形式(Lambers等人,2008;Zemunik et al., 2015)。虽然植物NAS随土壤养分有效性而变化,并影响植物群落组成(Johnson et al., 2023;Lambers et al., 2008;Li et al., 2021;Zemunik et al., 2015),它们在初级演替过程中影响植物物种更替的作用尚不清楚。植物NAS可以根据其获取不同营养形式的能力,即清除、挖掘和固定n2的策略,分为不同的类别(Lambers等,2008)。清除策略通过调节细根形态和与丛枝菌根(AM)、ericoid和外生菌根(ECM)真菌相关的共生关系,从土壤中获取植物可利用的养分。相比之下,采矿策略涉及动员和吸收不可用的营养物质,包括通过释放羧酸盐和质子来动员和吸收难溶的磷酸盐(如磷酸钙),以及通过释放水解酶来动员和吸收有机氮和磷。N2固定策略包括通过共生根瘤和根瘤菌固定大气中的N2。放线根植物一般能够通过丛状根系挖掘土壤中的磷,并通过根瘤菌获得氮(Shane &;琥珀,2005)。外生菌根植物可能同时采用这两种策略,因为ECM真菌延长菌丝来清除土壤养分,同时也通过ECM真菌或相关细菌分泌羧酸盐和酶来挖掘养分(landweert et al., 2001;袁等人,2024)。这些策略之间的权衡取决于土壤养分的可用性(Cao等,2024)。以往关于植物NAS与物种替代关系的研究主要是沿着数千年到数十万年的时间序列进行的(Holdaway et al., 2011;Zemunik et al., 2015)。例如,在澳大利亚西南部200万年的时间序列中,植物NAS的多样性被认为在植物群落组装中起着关键作用,因为P有效性在长期生态系统发展过程中下降(Zemunik et al., 2015)。然而,关于植物NAS对物种替代的长期贡献的知识并不一定能转移到几十年的快速和短期变化中。不同之处在于土壤养分动态在短期和长期时间序列之间的不同时间模式。土壤N和P的有效性通常下降,而顽固性有机养分在长期成土过程中随着土壤年龄的增加而积累(Turner &;Condron, 2013)。相比之下,在演替的早期阶段——从裸地到先锋群落的建立——土壤磷有效性和全氮含量在几十年内显著增加,主要受土壤磷形态变化和大气固氮作用的驱动(Zhou et al., 2013)。这些不同的养分有效性梯度和短期和长期时间序列之间不同的磷组成可能导致植物NAS在演替过程中对物种替代的贡献不同。然而,在土壤发生早期,植物NAS在物种替换中的作用在很大程度上被忽视了。在冰川后、海岸沙丘和火山年代序列中,土壤发育早期阶段土壤中氮的缺失已被广泛认可(lalibert<e:1> et al., 2012;Lawrence et al., 1967;Vitousek et al., 1993)。尽管年轻土壤含有丰富的全磷,而年老土壤则没有,这是由于土壤长期发育过程中的风化作用(Walker &;Syers, 1976),磷最近也被认为是这一早期阶段植物生长的限制因素(Darcy et al., 2018),因为不溶性磷灰石-磷主导土壤全磷,植物有效磷浓度非常低(Zhou et al., 2018)。由于采矿和固氮策略在贫瘠环境中获得稀缺可用营养物质的能力很强(Lambers, 2022;Reichert et al., 2022),可以推测具有固定n2和释放羧酸策略的植物物种比仅依赖清除策略的植物物种具有优势。 因此,具有采矿或固氮策略的物种经常在各种气候和母质的非常“年轻”的土壤群落中占主导地位(Chapin et al., 1994;Li et al., 2021;Vitousek et al., 1993;Zemunik et al., 2015;Zhou et al., 2013)。在这个阶段,外生菌根真菌可能释放更多的羧酸盐来挖掘磷,而较少依赖于远延伸菌丝的清除策略。这种对土壤养分有效性的反应可能有利于寄主植物物种,而不是严重依赖食腐策略的植物。相比之下,具有细根的植物物种和AM真菌获得足够的植物有效养分的能力较差。在原生演替初期至中期,土壤有效养分浓度增加,具有采掘策略的植物的表现也随之下降。在营养丰富的土壤中,清除策略更有效,因为与采矿策略相比,它们需要更少的碳投资(Raven等人,2018;Wang et al., 2022)。因此,利用食腐策略的植物可以更有效地获取植物可利用的营养物质,并获得竞争优势(Lambers等,2008)。例如,菌根植物可以更经济地获取磷,因此与那些通过释放羧酸盐来开采磷的植物相比,它们在富磷土壤中占主导地位(Zemunik等,2015)。此外,有限营养物质的获取与植物地上和地下光合作用的分配有关(Qiu et al., 2021)。在地下分配较少的光合作用以获取养分的植物在营养丰富的土壤中更受青睐(Tilman, 1985)。因此,与不同NAS和土壤养分状况相关的植物竞争力的剧烈变化可能是导致初生演替过程中优势种转移的原因之一。然而,支持这一论点的经验证据很少。为了解决这一知识空白,我们沿着青藏高原东部边缘海螺沟冰川前陆的原始植被演替的“年轻”时间序列(~130年)进行了研究(图S1)。在演替的前3个阶段,西藏棘(Elaeagnaceae)与白杨(Populus purdomii)在演替的第1阶段处于优势地位,在演替的第2阶段,两种物种共同处于优势地位。在此之后,purdomii逐渐取代西藏猿人成为第三阶段的优势种(Li &;熊,1995)。土壤氮浓度从几乎无法检测到的水平增加到相对较高的水平(Zhou et al., 2013)。在第一阶段,几乎所有的磷都存在于磷灰石或花岗岩中(Zhou et al., 2018)。然后随着磷灰石的快速风化和土壤年龄的增加,植物速效磷和有机磷浓度逐渐增加(Zhou et al., 2018,2019)。作为放线根植物类群,沙棘具有形成根状根系、簇状根系和AM共生的潜力(Oremus &;Otten, 1980,1981;Shah et al., 2015)。因此,Hippophae属的物种可以通过释放羧酸盐和质子来固定N2,动员不溶性P,通过分泌磷酸酶来水解有机P,并依靠AM真菌菌丝从土壤中清除植物可利用的N和P (Lambers等,2019)。另一方面,杨树属的物种表现出依赖于根外菌丝体的清除策略和利用共生真菌或相关细菌分泌的酶水解土壤中的有机氮和磷的挖掘策略(Phillips &;费伊,2006;袁等人,2024)。考虑到固氮和释放羧酸对N和P的获取效果,我们假设西藏杉树在初级演替开始时的优势地位是由于其固氮和采磷的能力比purdomii杉树更强。然而,通过包括细根和菌根在内的清除策略吸收养分可能随着植物有效土壤养分的增加而变得有效。因此,在演替过程中,purdomii代替西藏猿猴可能与后者在富营养化阶段比西藏猿猴的固氮和开采策略更有效有关。具体而言,我们旨在(1)研究两种植物NAS的差异和变化;(2)揭示植物NAS在初级演替过程中物种更替中的作用。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
